
Social Creativity - Induced Brain Activity Depending On Intelligence Components

Abstract
The authors identify the resting patterns of the frequency-spatial organization of cortical activity that associated with the verbal, visual-spatial, and social components of the intelligence. Taking into account the similarities between the development of verbal functions and that of social behavior, a model of their shared dynamic development was proposed. That is why the authors focused on finding out the specifics of relations between verbal, figural and social components of intelligence, optimal “pre-setting” of brain functional systems for implementing social creativity. For this purpose the following research methods - the Russian versions of the Amthauer and Gilford - Sullivan tests and EEG recording – were used. It was found that the original solution of a socially significant situation in the experimental model of creativity is accompanied by regionally specific synchronization of low- and high-frequency cortical oscillations. Having positive relations between the psychometrical indices of intelligence and creativity, the different EEG patterns correlate with these cognitive characteristics that indicate the availability of various strategies of search for the original idea depending on the individual profile of intelligence components. In conclusion, the authors note that the observed different frequently-spatial patterns of associations between cortical activation and intelligence components present the potential ways for correction of thinking strategies necessary for social creativity.
Keywords: Social creativitycortical activityEEG mapping
Introduction
The success of social adaptation, understanding of intentions and behavior of other people allows «social intelligence» by the functions of “social brain” (Abu-Akel & Shamay-Tsoory, 2011; Adolphs, 2003; Valk et al., 2017). The “Theory of Mind” (ТоМ) (Adolphs, 2003; Bar-On, Tranel, Denburg, & Bechara,, 2003; Baron-Cohen et al., 1999) is proposed for describing neurobiological mechanisms behind the organization of social behavior. According to ToM the understanding of other people behavior which is necessary for their personal social adaptation develops within the first three years and includes the same functions as those which are necessary for language development. Taking into account the similarities between the development of verbal functions and that of social behavior, a model of their shared dynamic development was proposed. The understanding of other people intentions facilitates language development at its early stages, and then language acquisition and verbal intelligence greatly influence the improvement of social intelligence (Malle, 1999). The means of language activity needed for successful development of social intelligence are the following: vocabulary including the words which denote emotional reactions and states, the abilities to identify and use the intonation and emotional prosodies. Psychometrically, the verbal intelligence consists in the abilities to understand and make use of various speech operations.
In addition to speech functions, the development of social adaptation requires the improvement of visual and spatial abilities including the understanding of information when perceiving facial expressions and gestures in the existing situations.
Visual perception of eyes and mouth positions, the analysis of body position and movements are provided by the interaction of neuronal structures (inferior parietal lobule and medial cortical structures) representing the mirror neuron system and metalizing system (Adolphs, 2003; Overwalle & Baetens, 2009). In its turn, the speed and effectiveness of information processing of visual modality at the psychometrical level is determined by the measure of visual-spatial intelligence. Thus, both visual and verbal abilities are indispensable for social intelligence development.
There is a view that social intelligence can be presented by three components: social sensitivity reflecting the conclusions of social roles and social contacts, social insight including the understanding of social and psychological conditions and moral assessment of events, and social communication (Greenspan & Wieder, 1997). The Structure of Intellect Theory proposed by J. P. Guilford included divergent and convergent thinking and found its application in M. Sullivan’s techniques. It deals with the measure of social intelligence, the definition of abilities to interpret people’s mental state and the results of their behavior (based on convergent thinking), as well as the evaluation of flexibility of knowledge about changing social behavior and abilities to predict possible outcomes in interpersonal situations (divergent functions).
All this laid the foundation for understanding the creative social intelligence, which includes the divergent means of assessment, relationships, transformations, and predicting people’s interactions (Hendricks, Guilford, & Hoepfner, 1969). Consequently, it is possible to determine two different parts of social intelligence: the understanding of people’s behavior on the basis of existing socio-cultural stereotypes and various forms of social adaptation based on divergent interpretation of the content of behavior.
Social creativity is the necessary component of modern innovative activity which requires new forms of leadership and interactions between team members (Fischer & Shipman, 2011; Hahn, Kun, & Choi, 2011).
Various models are used as neurophysiologic mechanisms of organizing social behavior along with the Theory of Mind (ToM). One of them is a dual system, its function is to coordinate conscious control of information processing and decision making by top-down processing and automatic bottom-up processing (Spunt, 2015). The second is C (reflective) /X (reflexive) system, proposed for unconscious and automatic gaining of social experience or targeted social learning (Lieberman, 2007). The intuitive social behavior or intuitive social thinking are formed on the basis of implicit learning and non-verbal coding of information entering the mind in order to form various life scenarios and rules of behavior within the framework of various social roles. The C-system of social behavior regulation allows quick orientation in new situations taking into account the acquired experience and instructions on activity control. The central structures of this system are medial temporal lobe, posterior parietal lobe and lateral prefrontal cortex.
Focusing on psychometrical assessment of social intelligence or verbal or non-verbal creativity, one might assume that for determining the abilities of social creativity, the stimuli reflecting various life situations can be used. Taking this hypothesis into consideration, we have developed the techniques for determining social creativity in experimental conditions (Finikov & Razumnikova, 2009). Two variants have been formed on the basis of previous expert assessment of stimulus material: in the first variant, social roles and interaction of subjects are viewed in everyday situations; in the second one, the situation is quite paradoxical. In the first variant of determining social creativity (i.e. convergent problem), the initial adequacy and the speed of understanding a social situation appear in the fore, and only the flexibility of social thinking allows moving away from the stereotype and seeking for other solutions of the problem. The second variant is intended to stimulating the divergent thinking. In this case, the main difficulty is not to overcome the stereotypes but to understand the problem itself and be motivated to finding various interpretations of the situation under investigation.
Problem Statement
The study of frequency-spatial organization of brain activity using the developed techniques for measuring social creativity has shown that the solution of socially important problems in experimental conditions is accompanied by power increase of the delta and gamma 2 rhythms and desynchronization of the alpha 2 oscillations. When solving the problems of a divergent character, this increase is, however, less-pronounced than it is at interpreting the “convergent” situation (Razumnikova & Finikov, 2010).
This effect of less-pronounced brain activation in divergent solution of a social problem corresponds to the data which show that creative thinking based on the defocused or internal attention is characterized by comparatively weak brain activation at alpha rhythm frequencies (Fink, Grabner, Benedek, & Neubauer, 2006; Martindale, 1999). The obtained results can be interpreted as the evidence of automatic and intuitive mobilization of memory resources, which keep implicitly-formed understanding of social roles of the subjects according to certain experimental situations. This hypothesis of “insight” i.e., a suddenly appearing solution of social creative problems, is supported by the relationship between the gamma rhythm and insight strategy of thinking, which was discovered earlier at testing verbal creativity (Jung-Beeman et al., 2004).
Desynchronization of alpha2 biopotentials induced by task performance is a result of summation of activation and inhibition processes in the cortex with the latter dominating when the internal attention is increased as the changes of alpha rhythm amplitude reflect the specify of free-oriented or targeted selection of visual information (Rihs, Michel, & Thut, 2007), which is necessary for finding answers while interpreting socially variable information.
The originality of presented situation interpretation appeared to be related with a background cortex activity at low frequencies of delta rhythm and at high frequencies of gamma rhythm, which reflects presumably endogenous components of social thinking: motivation of solving the presented problem and individual characteristics of selecting visually presented information (Razumnikova & Finikov, 2011). This positive correlation between the originality and the power of rhythms is presented in posterior areas of the right hemisphere while performing the “divergent” task. The obtained patterns for delta- and gamma-activity as the predictors of social creativity are to be considered as EEG-correlates of summation of endo - and exogenous components of social thinking, in this case, neuron systems of frontal cortex dominate in formation of endogenous component of social creativity whereas the posterior areas of cortex – in that of exogenous component of social creativity.
Widespread gamma2 oscillations as a predictor of answer fluency reflect the possibility of generating a variety of ideas that can be both original and stereotyped solutions of social problems, the creation of these ideas requires different information resources of the brain.
The latest intensive research of networks of spontaneous activity in the brain (resting state networks (RSN) or default mode network – (DMN)) is due to the fact that this activity reflecting individual characteristics of subsequent functional information processing forms different strategies of
Research Questions
How are the different components of intelligence and social creativity interrelated? How are these components associated with oscillatory brain activity? Which are the patterns of brain oscillations as predictors of both social creativity and different components of intelligence?
Purpose of the Study
Thus, the aim of our research is to find out the specificity of relationships between verbal, figural and social components of intelligence, which could be considered as optimal “pre-setting” of brain functional systems for implementing social creativity.
Research Methods
Participants
Forty two right-handed students participated voluntary in the research (aged between 17 and 20 years, 18 of them are men). This study was approved by the local Ethics Committee of both Novosibirsk State Technical University, and Research Institute of Physiology and Basic Medicine. All participants signed an informed consent form before the experiment.
Assessment of intelligence and creativity
In order to measure verbal, figural and social components of intelligence, the Russian versions of the Amthauer and Gilford - Sullivan tests were used. The intelligence test was performed for two to three weeks, which preceded the EEG records. Verbal (IQv), visual-spatial (IQf) and social (IQs) components of intelligence were considered in statistic data analysis. In order to find the indices of social originality (answers obtained at recording EEG in the experimental model of social creativity (Razumnikova & Finikov, 2010) a previously-developed computerized method was used, according to which the index of originality is calculated as the number that is inverse to the number of similar ideas kept in previously created database (Finikov & Razumnikova, 2009).
EEG recording
The EEG was recorded unipolarly by means of Ag/AgCl electrodes located in an electrode ca[ in 19 positions (Fp1, Fp2, F7, F3, Fz, F4, F8, T7, C3, Cz, C4, T8, P7, P3, Pz, P4, P8, O1, O2) with the use of the encephalograph “Neurovisor BMM40” (“Neurobotics”, Moscow). The joint ear electrode was used as a reference one. The EEG signals were sampled at a frequency of 1000 Hz, the filters of low and high frequencies are 0.5 and 70 Hz, respectively. Analog-to-digital conversion of the signal and calculation of power distribution parameters of EEG with the use of The were performed on the basis of the “Neurocortex4.4” software.
The power of biopotentials is calculated for nine frequency ranges: delta (0.5–4 Hz), theta1 (4–6 Hz), theta 2 (6–8 Hz), alpha1 (8–10 Hz), alpha2 (10–13 Hz), beta1 (13–20 Hz), beta2 (20–30 Hz), gamma1 (30–40 Hz) и gamma 2 (40–50 Hz).
According to the aim of research, in analyzing the data, power indices of biopotentials in the background (with closed eyes) were used. These indices were considered in relation to intelligence and social creativity components, the latter being obtained during the test of its EEG correlates.
Findings
Behavioral data
When analyzing the intelligence and creativity, we found positive correlation between originality (Ors) and fluency (Fls) indices according to psychometric assessment of social originality (р<0.0001) and between various components IQ (Table
EEG mapping data
At the first step, we analyzed the association of social creativity and intelligence components with total power of oscillations in 9 frequency bands. For different components IQ, the significant relationship was found for low frequency delta band and high frequency beta2 band. In bands theta1 and gamma2, only the tendencies of correlation IQ and biopotential power (Table
The next step of this analysis was aimed at revealing regional specificity of EEG patterns in the background state associated with verbal (IQv), visual-spatial (IQf) and social (IQs) components of intelligence and/or social creativity. We analyzed only those frequency bands which had been identified at the first step of analysis.
For the delta band, the positive correlations Ors и Fls with the right hemisphere oscillations were found in F8 and T8 (0.01<p<0.1) (Fig.1). According to the results of regressive analysis, the reliable predictor Ors is the power of delta rhythm in F8, which allows the description of 21% of variability Ors (Table
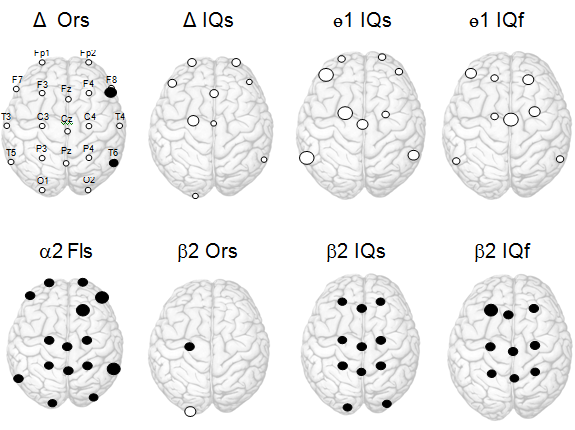
Ors – originality, Fls – fluency, IQs – social intelligence, IQf – visual-spatial intelligence, ● - positive, ○ - negative correlation, size of circles indicate 0.001<p<0.1
The analysis of IQ components showed that the negative relationships of the delta rhythm were characteristic of only IQs: they were widely spread over the region, but they were more pronounced for the anterior cortex at left hemisphere domination (Fig.1). According to the results of regression analysis, the predictor IQs is the delta rhythm in C3, which accounts for 17% of its variability (Table
In theta1 band, Ors was positively related to the power of oscillations in F8 and T8 (p<0.04), however, the reliable predictor was only the rhythm power in T8, which explained about 18% of creativity index dispersion. Negative relationships between IQs and theta oscillations were reliable for leads F7, T7, C3, C4 (0.004<p<0.02), but the reliable predictor IQs was the power theta in C3 (Table
The alpha 2 band was characterized by the generalized distributed correlations Fls and alpha oscillations (Fig.1), but only for T8 there was a reliable relationship between the power alpha 2 and Ors, and this cortical area according to the regression model of creativity indexes was essential for explaining the variations of both Ors and Fls (Table
Ors and Fls had similar regression models, which included high frequency beta 2 oscillations: a higher level of social creativity was characterized by the increase of the rhythm power in С3 and the decrease in О1 (24%<R2 <28%). IQs and IQf are positively related to the beta 2 rhythm power, which is generally presented in the frontal and parietal cortex (Fig.1), however, the significant predictors of various components of intelligence proved be locally presented beta2 oscillations: for IQs – in Fz и O1, for IQf – in Cz, and for IQv – in Fz (10%<R2 <21%, Table
At the frequency of the gamma2 biopotentials, 13% of Ors and Fls variability and 10% of IQf were related to the rhythm power in О1, but of different signs, and 12% of IQs – to the power of gamma oscillations in Fz (Table
Thus, though the psychometrically assessed components of intelligence and creativity are closely related (see Table
The discovered positive relationships between alpha 2 rhythm and creativity indices are consistent with the hypothesis about the increase of internal attention as a condition of successful problem solving based on divergent thinking (Fink et al., 2006). The maximum significance of this presetting of posterior temporal areas of the right hemisphere is in good agreement with the views about intuitive use of resources of implicitly formed memory (Jung-Beeman et al., 2004), which allows different interpretation of social situations being modeled in the experiments.
It is notable that the contribution of alpha2 rhythm to IQf is unidirectional with Fls and Ors and marked for the central prefrontal cortex that can be interpreted as a preparation for involving the executive control of attention while solving a problem. A more pronounced association of the resting brain activity with IQf, presented at different frequencies along the central line: Fz, Cz and, at last, О1, but not with IQv. The effect can be explained by the fact that verbal functions, on the one hand, are widely spread over the systems of brain (Friederici, 2011; Skeide, Brauer, & Friederici, 2016), and, on the other hand, they are automated very largely, this is perhaps why the specialization of IQv is lost in averaging the background EEG, and it is found only in Fz at the beta2 frequency.
The steady informative effect of loci Fz, Cz, Рz, characterizing IQs и IQf (see Figure
Beta2 oscillations are related to both speech processes and various forms of selecting information which is represented in locally organized neural networks (Engel & Fries, 2010). The predictors of all the considered IQ components, which have been obtained at this frequency, are indicative of their congruent-to-creativity relationship with preparing the DMN system for operation. Negative correlation between Ors, Fls and beta2 synchronization in the left occipital cortex indicates that low background activity can facilitate a further generation of visual images while testing social creativity. The discovered relationship between creativity, intelligence and gamma activity is in line of this hypothesis.
EEG correlates of social creativity are presented more locally in comparison with IQ, as they are the efficiency indices of social situation interpretation only in one experimental model. In this case, the right hemisphere foci of background brain activity (Т8) in the wide frequency band from delta to alpha2 can be considered as an evidence of the presetting of brain activity which accompanies further domination of the right hemisphere in search for original solution of creative tasks including social content (Kounios et al., 2008; Razumnikova & Finikov, 2010, 2011).
Conclusion
Social creativity as the ability to original and flexible solution of socially significant problems is reflected in the specificity of hemispheric activity of the brain and depends on the functional state of the DMN system due to the individual structure of intellectual abilities.
The observed different frequently-spatial patterns of associations between cortical activation and intelligence components present the potential ways for correction of thinking strategies necessary for social creativity.
Acknowledgments
The authors wish to thank Semen Finikov for EEG registration.
References
- Abu-Akel, A., & Shamay-Tsoory, S. (2011). Neuroanatomical and neurochemical bases of theory of mind. Neuropsychologia, 49(11), 2971-2984.
- Adolphs, R. (2003). Cognitive neuroscience of human social behavior. Nat. Rev. Neurosci., 4 (3), pp. 165-178.
- Bar-On, R., Tranel, D., Denburg, N. L., & Bechara, A. (2003). Exploring the neurological substrate of emotional and social intelligence. Brain, 126, 1790-1800.
- Baron-Cohen, S., Ring, H. A., Wheelwright, S., Bullmore, E. T., Brammer, M., Simmons, A., … & Williams, S. (1999). Social intelligence in the normal and autistic brain: an fMRI study. Eur. J. Neurosci., 11, 1891-1898.
- Cabral, J., Luckhoo, H., Woolrich, M., Joensson, M., Mohseni, H., … & Deco, G. (2014). Exploring mechanisms of spontaneous functional connectivity in MEG: how delayed network interactions lead to structured amplitude envelopes of band-pass filtered oscillations. Neuroimage, 90, 423-35.
- Deco, G., & Corbetta, M. (2011). The dynamical balance of the brain at rest. Neuroscientist, 17(1), 107-123.
- Engel, A. K.,& Fries, P. (2010). Beta-band oscillations – signaling the status quo? Current Opinion in Neurobiology, 20 (2), 156-165.
- Finikov, S. B., & Razumnikova, O. M. (2009). Development of a method for quantitative assessment of social creativity in experimental conditions. Bull. SB RAMS, 5 (139), 68-71. [in Rus.].
- Fink, A., Grabner, R. H., Benedek, M., & Neubauer A. C. (2006). Divergent thinking training is related to frontal electroencephalogram alpha synchronization. Eur. J.Neurosci, 23, 2241–2246.
- Fischer, G., & Shipman, F. (2011). Collaborative design rationale and social creativity in cultures of participation. Human Technology, 7(2), 164-187.
- Friederici, A. D. (2011). The brain basis of language processing: from structure to function. Physiol Rev., 91, 1357–1392.
- Greenspan, S. I., & Wieder, S. (1997). Developmental patterns and outcomes in infants and children with disorders in relating and communicating: A chart review of 200 cases of children with autistic spectrum diagnoses. J. Developmental and Learning Disorders, 1, 87-141.
- Hahn, M. H., Kun, C. L., & Choi, D. Y. (2011). Effects of social and emotional intelligence on the creative process and individual creativity. In: U-and E-Service, Science and Technology. Berlin, Heidelberg: Springer.
- Harmony, T. (2013). The functional significance of delta oscillations in cognitive processing. Front Integr Neurosci., 7, 83.
- Hearne, L. J., Mattingley, J. B., & Cocchi, L. (2016). Functional brain networks related to individual differences in human intelligence at rest. Scientific Reports, 6, 32328.
- Hendricks, M., Guilford, J. P., & Hoepfner, R. (1969). Measuring creative social intelligence. Reports from the Psychological Laboratory. Univ. Southern California.
- Huster, R. J., Enriquez-Geppert, S., Lavallee, C. F., Falkenstein, M., & Herrmann, C. S. (2013). Electroencephalography of response inhibition tasks: functional networks and cognitive contributions. Int J Psychophysiol., 87(3), 217-233.
- Jung-Beeman, M., Bowden, E. M., Haberman, J., Frymiare, J. L., Arambel-Liu, S., Greenblatt, R., Reber, P.J., & Konnios J. (2004). Neural activity when people solve verbal problems with insight. PLoS Biol, 2 (4), 0500–0510.
- Jung, R. E., & Haier, R. (2007). The Parieto-Frontal Integration Theory (P-FIT) of intelligence: Converging neuroimaging evidence. Behavioral and Brain Sciences, 30(2), 135-154.
- Knyazev, G. G. (2012). EEG delta oscillations as a correlate of basic homeostatic and motivational processes. Neuroscience and Biobehavioral Reviews, 36, 677–695.
- Kounios, J., Fleck, J. I., Green, D. L., Payne, L., Stevenson, J. L., Bowden, E. M., & Jung-Beeman, M. (2008). The origins of insight in resting-state brain activity. Neuropsychologia, 46(1), 281-291.
- Lieberman, M. D. (2007). Social cognitive neuroscience: A review of core processes. Annu. Rev. Psychol., 58, 259-289.
- Malle, B. F. (1999). How people explain behavior: A new theoretical framework. Personality and Social Psychology Review, 3, 23-48.
- Martindale, C. (1999). Biological bases of creativity. In: Handbook of Creativity. Ed. Sternberg R.J. Cambridge: Cambridge Univ. Press. pp. 137–152.
- Overwalle, F. van., & Baetens, K. (2009). Understanding others' actions and goals by mirror and mentalizing systems: a meta-analysis. Neuroimage, 48(3), 564-584.
- Razumnikova, O. M., & Finikov, S. B. (2010). EEG correlates of social creativity. Zh Vyssh Nerv Deiat Im I P Pavlova, 60(6), 691-695. [in Rus.].
- Razumnikova, O. M., & Finikov, S. B. (2011). Topographic characteristics of cortex activity in delta-, alpha2-, and gamma2- frequency bands related to social creativity. Zh Vyssh Nerv Deiat Im I P Pavlova. 61(6), 706-715. [in Rus.]
- Rihs, T., Michel, C. M., & Thut, G. (2007). Mechanisms of selective inhibition in visual spatial attention are indexed by alpha band EEG synchronization. Eur. J. Neurosci., 25, 603–610.
- Robinson, D. L., & Behbehani, J. (1997). Intelligence differences: neural transmission errors or cerebral arousability? Kybernetes, 26, 407-424.
- Skeide, M. A., Brauer, J., & Friederici, A. D. (2016). Brain functional and structural predictors of language performance. Cerebral Cortex, 26, 2127-2139.
- Spunt, R. P. (2015). Dual-process theories in social cognitive neuroscience. Brain Mapping: An Encyclopedic Reference, 3, 211-215.
- Takeda, Y., Hiroe, N., Yamashita, O., & Sato, M. (2016). Estimating repetitive spatiotemporal patterns from resting-state brain activity data. NeuroImage, 133, 251–265.
- Valk, S. L., Bernhardt, B .C., Trautwein, F.-V., Böckler, A., Kanske, P., Guizard, N., … & Singer, T. (2017). Structural plasticity of the social brain: Differential change after socio-affective and cognitive mental training. Sci. Adv., 3(10), e1700489.
Copyright information

This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.
About this article
Publication Date
12 December 2019
Article Doi
eBook ISBN
978-1-80296-073-0
Publisher
Future Academy
Volume
74
Print ISBN (optional)
-
Edition Number
1st Edition
Pages
1-419
Subjects
Society, culture, education
Cite this article as:
Pusikova, M. V., & Razumnikova*, O. M. (2019). Social Creativity - Induced Brain Activity Depending On Intelligence Components. In S. Ivanova, & I. Elkina (Eds.), Cognitive - Social, and Behavioural Sciences - icCSBs 2019, vol 74. European Proceedings of Social and Behavioural Sciences (pp. 365-375). Future Academy. https://doi.org/10.15405/epsbs.2019.12.02.43